Glycogen Metabolism
Glycogen is a storage form of glucose. It is a large, branched polymer of glucose residues that can be broken down to yield glucose molecules when energy is needed. Unlike, fatty acids, which are more reduced than glycogen and thus contain more energy, glucose can provide energy in the absence of oxygen and can thus supply energy for anaerobic activity. The two major sites of glycogen storage are the liver and skeletal muscle, but more glycogen is stored in skeletal muscle overall because of muscle's much greater mass. Glycogen is present in the cytoplasm, and in liver cells, glycogen synthesis and degradation are regulated to maintain blood glucose levels as required to meet the needs of the organism as a whole. In contrast, in muscle these processes are regulated to meet the energy needs of the muscle itself.
Glycogen Degradation Glycogen degradation consists of three steps: (1) release of glucose-1-phosphate from glycogen, (2) remodeling of glycogen substrate to permit further degradation, and (3) the conversion of glucose-1-phosphate into glucose-6-phosphate for further metabolism. | Glycogen Synthesis Glycogen synthesis requires an activated form of glucose, (UDP-glucose), which is formed by the reaction of UTP and glucose-1-phosphate. UDP-glucose is added to nonreducing ends of glycogen, and the molecule must be remodeled for continued synthesis. |
Regulation of Glycogen Synthesis and Degradation
Several enzymes taking part in glycogen metabolism allosterically respond to metabolites that signal the energy needs of the cell. Through these allosteric responses, enzyme activity is adjusted to meet the needs of the cell.
Hormones may also initiate signal cascades that lead to the reversible phosphorylation of enzymes, which alters their catalytics rates. Regulation by hormones adjusts glycogen metabolism to meet the needs of the organism.
Enzymes of Glycogen Breakdown
The breakdown of glycogen requires four enzymes: one to degrade glycogen, two to remodel glycogen so that it remains a substrate for degradation, and one to convert the product of glycogen breakdown into a form that can be used for further metabolism.
Glycogen Phosphorylase is the key enzyme in glycogen breakdown. Glycogen phosphorylase cleaves glycogen by the addition of Pi to yield glucose-1-phosphate. The phosphorylytic cleavage of glycogen is energetically advantageous because they released sugar is already phosphorylated. Also, muscle cells do not have transporters for glucose-1-phosphate, therefore whatever glycogen is broken down in the muscle cell stays in the muscle cell.
Debranching enzyme is also needed for the breakdown of glycogen. The alpha-1,6-glycosidic bonds at the branch points are not susceptible to cleavage by phosphorylase. When phosphorylase reaches a terminal residue four residues away from a branch point, it stops cleaving the 1,4-bonds. Two additional enzymes, a transferase and alpha-1,6-glucosidase, remodel the glycogen for continued degradation by the phosphorylase. With four residues left on each branch, the transferase shifts a block of three glucosyl residues from one outer branch to the other. Thus, one glucose residue is left, joined by an alpha-1,6 bond. alpha-1,6-glucosidase, the true debranching enzyme, hydrolyzes the alpha-1,6 bond, releasing a free glucose molecule. The free glucose is phosphorylated by hexokinase (glycolytic enzyme).
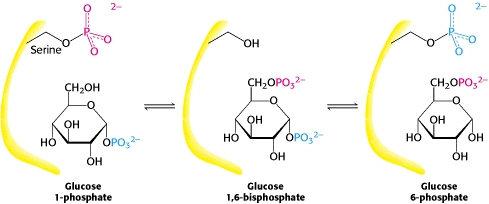
Phosphoglucomutase converts glucose-1-phosphate to glucose-6-phosphate, such that the glucose can enter the metabolic mainstream. A phosphoryl group is exchanged between the enzyme and substrate. Note: the same phosphoryl group is not just moved.

|
phosphoglucomutase.jpg Size : 0.038 Kb Type : jpg |
{kind=link}
phosphoglucomutase.jpg (c) 2007 W.H. Freeman and Company
Glucose-6-Phosphatase: in liver only
A major function of the liver is to maintain a nearly constant level of glucose in the blood. The liver releases glucose during muscle activity and between meals, and the released glucose is taken up by the brain and skeletal muscle. The phosphorylated glucose produced from glycogen metabolism, however, is not transported out of cells. Therefore, the liver contains glucose-6-phosphatase, a hydrolytic enzyme, that enables glucose to leave the organ. Glucose-6-phosphatase is present on the lumenal side of the smooth endoplasmic reticulum membrane. Glucose-6-phosphatase is not present in most other tissues, therefore, these tissues retain glucose-6-phosphate for the generation of ATP. Glucose is not a major fuel for the liver, therefore, it can be transported out of the liver with relatively few consequences.
Regulation of Glycogen Phosphorylase
Glycogen phosphorylase is regulated by several allosteric effectors that signal the energy state of the cell as well as by reversible phosphorylation, which is responsive to hormones such as insulin, epinephrine, and glucagon. The muscle uses glucose to produce energy for itself, but the liver maintains glucose homeostasis of the organism as a whole.
Muscle phosphorylase is regulated by the intracellular energy charge. Muscle phosphorylase b is active only in the presence of high concentrations of AMP, which acts as an allosteric effector by binding to a nucleotide binding site and stabilizing the conformation of phosphorylase b in the active state. ATP acts as a negative allosteric effector by competing with AMP. Glucose-6-phosphate also favors the less active state of phosphorylase b (i.e. feedback inhibition). Phosphorylase b is converted to phosphorylase a by phosphorylation of a serine residue in each subunit. Epinephrine will trigger phosphorylation of phosphorylase b through the action of phosphorylase kinase. The absence of glucose-6-phosphatase is muscle ensures that the glucose-6-phosphate released from glycogen remains within the cell.
Liver phosphorylase produces glucose for use by other tissues. The role of glycogen degradation in the liver is to produce glucose that is capable of being exported to other tissues when blood glucose is low. Liver phosphorylase is sensitive to the presence of glucose; the binding of glucose deactivates the enzyme. Unlike muscle phosphorylase, liver phosphorylase a is the most responsive R-to-T transition, and also, liver phosphorylase is insensitive to regulation by AMP.
Phosphorylase kinase is the enzyme that activates phosphorylase b by attaching a phosphoryl group. The kinase is activated both by phosphorylation and by increases in calcium levels. Phosphorylase kinase in coverted from a low-activity form into a high activity one by phosphorylation of its beta subunit. Phosphorylase kinase can also be partly activated by high calcium levels. The delta subunit of phosphorylase kinase is calmodulin, a calcium sensor that stimulates many enzymes in eukaryotes. Note: activation by calcium becomes important in muscle cells, where contraction is triggered by calcium release. Phosphorylase kinase attains maximum activity only when it is phosphorylated and calcium is bound.
Hormonal Signals for Glycogen Breakdown
Protein kinase A activates phosphorylase kinase, which in turn activates glycogen phosphorylase. Glucagon and epinephrine trigger breakdown of glycogen. Epinephrine stimulates glycogen breakdown in muscle, and, to a lesser extent, in the liver. However, the liver is more responsive to glucagon, which is secreted by the alpha cells of the pancreas when blood glucose levels are low.
Epinephrine and glucagon initiate a signal-transduction cAMP cascade
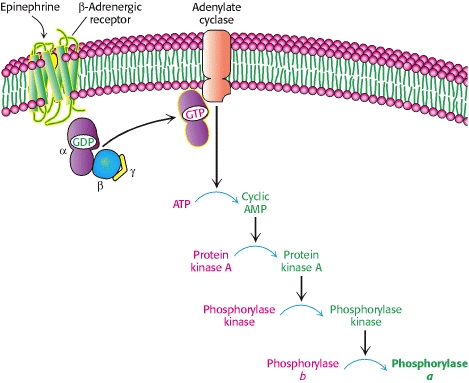
Recall: epinephrine will bind to beta-2 adrenergic receptors in skeletal muscle and glucagon will bind to glucagon receptors in the liver.
Note: epinephrine can also elicit glycogen degradation in the liver. In addition to binding to beta-AR, it binds alpha-AR, which initiates the phosphoinositide cascade that induces calcium release. Remember also, phosphorylase kinase contains a calmodulin subunit.
When glucose needs have been satisfied, phosphorylase kinase and glycogen phosphorylase are dephosphorylated and inactivated, meanwhile glycogen synthesis is activated. When the initiating hormone is no longer present, the glycogen phosphorylase is shut down. Protein kinase A sets the stage for the shutdown of glycogen degradation by adding a phosphoryl group to the alpha-subunit of phosphorylase kinase. The addition of another phosphoryl group make the phosphorylase kinase a better substrate for dephosphorylation by protein phosphatase I, which removes the phosphoryl group from glycogen phosphorylase, converting it into the inactive b form.
Glycogen Synthesis
Glycogen synthesis uses uridine diphosphate glucose (UDP-glucose) as an activated donor, rather than glucose-1-phosphate. UDP-glucose is an activated form of glucose. The C-1 of a glucosyl unit is attached to the nucleotide diphosphate moeity. UDP-glucose is synthesized from glucose-1-phosphate and UTP in a reaction catalyzed by UDP-glucose pyrophosphorylase, liberating the outer two phosphoryl residues of UTP as pyrophosphate (PPi). PPi is rapidly hydrolyzed to Pi + Pi making the reaction essentially irreversible. The activated glucosyl unit of UDP-glucose is transferred to the hydroxyl group of the C-4 of the terminal residue to form an alpha-1,4-glycosidic bond. UDP is displaced in this reaction catalyzed by glycogen synthase, which is also the key regulatory enzyme of glycogen synthesis. Glycogen synthesis requires a primer of 4 residues, this function is carried out by glycogenin, which is made up of two subunits, each of which catalyzes the addition of eight glucose residues to the other subunit. Glycogen synthase then takes over to extend the molecule.
A branching enzyme form the alpha-1,6-linkages. A branch is created by breaking an alpha-1,4 linkage and forming and alpha-1,6 linkage. A block of 7 residues is taken from a chain of at least 11 residues and transferred to the new branch point, which must be at least 4 residues away from a preexisting branch. Branching increases the solubility of glycogen, and creates a large number of non reducing ends, thus increasing the rates of synthesis and degradation.
Regulation of Glycogen Synthase
Glycogen synthase is regulated by covalent modification. Glycogen synthase is phosphorylated at multiple sites by multiple enzymes, notably, protein kinase A and glycogen synthase kinase. Phosphorylation converts the active a form of glycogen synthase into the inactive b form. The phosphorylated b form is active only in the presence of high concentrations of glucose-6-phosphate, and allosteric activator.
Glycogen Synthesis and Degradation are Reciprocally Regulated
The glucagon-epinephrine cAMP cascade that triggers glycogen degradation in liver and muscle, also shuts off glycogen synthesis. Glucagon and epinephrine control both synthesis and degradation through protein kinase A. Protein phosphatase I reverses the regulatory effects of kinases on glycogen metabolism. When glycogen replenishment is needed, such as after exercise, the first step is to shut down the phosphorylated proteins that stimulate glycogen breakdown. Protein phosphatases catalyze the hydrolysis of phosphorylated serine and threonine residues in proteins. Protein phosphatase I inactivates glycogen phosphorylase by dephosphorylation, thereby reversing the effects of the phosphorylation cascade. Moreover, protein phosphatase I removes phosphoryl groups glycogen synthase b to convert it to the a form. When glycogen degradation is called for, protein kinase A reduces the activity of protein phosphatase I. In muscle, the Gm regulatory subunit of protein phosphatase I will be phosphorylated, and the catalytic subunit is released from glycogen and its substrates. Also, almost all tissues contain small proteins that, when phosphorylated, bind to the catalytic subunit of protein phosphatase I and inhibit it.
Actions of Insulin in Glycogen Metabolism
When blood glucose is high, insulin stimulates the synthesis of glycogen by inactivating glycogen synthase kinase. Insulin binds to the tyrosine kinase receptor in the plasma membrane, activating the activity of the tyrosine kinase, so that it phosphorylates insulin-receptor substrates, which trigger signal transduction pathways that ultimately lead to the activation of protein kinases that phosphorylate and inactivate glycogen synthase kinase. Thus protein phosphatase I can dephosphorylate glycogen synthase.
Liver Regulates Blood Glucose Levels
Insulin is the primary signal for glycogen synthesis, but plasma glucose concentration can also be used. Phosphorylase a in liver cells is the glucose sensor. The binding of glucose to phosphorylase a causes a transition from R to T for glycogen phosphorylase. The change from R to T renders the phosphoryl group a substrate for protein phosphatase I. PPI binds tightly to phosphorylase a in the R form, but is released and active upon phosphorylase transition to the T state. Phosphorylase b does not bind PPI and thus conversion of phosphorylase a to phosphorylase b releases PPI, which is then free to activate glycogen synthase.
Berg, J., Tymoczko, J., Stryer, L. Biochemistry 6E. ©2007 W.H. Freeman and Company