Contrasting Fatty Acid Synthesis and Degradation
(1) Synthesis takes place in the cytoplasm, degradation takes place in the mitochondrial matrix
(2) Intermediates in fatty acid synthesis are covalently linked to acyl-carrier protein (ACP), whereas intermediates in fatty acid breakdown are covalently attached to the sulfhydryl group of CoA
(3) The enzymes of fatty acid synthesis are joined in a single polypeptide chain
(4) Malonyl ACP donates two carbon units for the addition to the growing fatty acid chain. Elongation is driven by the release of carbon dioxide.
(5) The reductant in fatty acid synthesis is NADPH
(6) Elongation stops on the formation of palmitate (C-16).
Malonyl CoA
The formation of malonyl CoA is the committed step in fatty acid synthesis. Acetyl CoA is carboxylated by acetyl CoA carboxylase to form malonyl CoA and one ATP is used. Acetyl CoA carboxylase contains a biotin prosthetic group.
Acyl Carrier Protein
The intermediates in fatty acid synthesis are linked to an acyl carrier protein.
Fatty Acid Synthesis
Fatty acid synthase is a complex of distinct enzymes that catalyze the synthesis of long chain fatty acids.
The elongation phase of fatty acid synthesis begins with the formation of acetyl-ACP and malonyl-ACP. These reactions are catalyzed by acetyl transacylase and malonyl transacylase respectively. (Note: malonyl CoA is formed by the carboxylation of acetyl CoA at the expense of 1 ATP).
Acetyl-ACP and malonyl-ACP react to form acetoacetyl-ACP, catalyzed by the acyl malonyl ACP condensing enzyme. Carbon dioxide is released and thus a four carbon unit is formed.
Acetoacetyl ACP is then reduced to D-3-hydroxybutyryl ACP and NADPH is oxidized to NADP+. This reaction is followed by a dehydration followed by another reduction to butyryl ACP, NADPH is again oxidized.
Thioesterase cleaves ACP from the fatty acid chain at the end of synthesis.
Transfer of Acetyl CoA to Cytoplasm
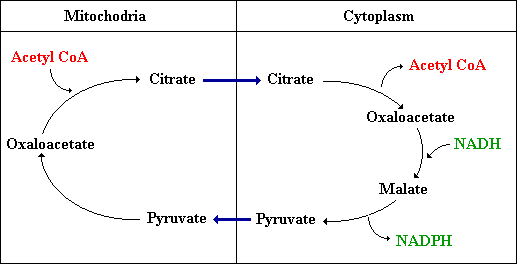
Recall, fatty acids are synthesized in the cytoplasm from acetyl CoA, but acetyl CoA is formed in the mitochondria from pyruvate. Acetyl CoA cannot cross the inner membrane therefore there must be a transport mechanism. Citrate carries acetyl groups across the inner mitochondrial membrane. Citrate is formed in the mitochondrial matrix by the condensation of acetyl CoA with oxaloacetate, and when present at high levels, citrate is transported to the cytoplasm and cleaved by ATP-citrate lyase. Thus, acetyl CoA and oxaloacetate are transferred from matrix to cytoplasm at the expense of one ATP.
Sources of NADPH
Oxaloacetate that was transported to the cytoplasm with acetyl CoA must be transported back to the matrix. The following reactions are important because they generate NADPH.
(1) oxaloacetate is reduced to malate by NADH by malate dehydrogenase
(2) malate is oxidatively decarboxylated by an NADP+-linked malate enzyme, which reduces a molecule of NADP+ to NADPH forming pyruvate.
(3) pyruvate readily enters the mitochondria, where it is carboxylated to oxaloacetate by pyruvate carboxylase.
Thus, one molecule of NADPH is generated for each molecule of acetyl CoA that is transferred from mitochondria to cytoplasm. Therefore, for the synthesis of palmitate, eight molecules of NADPH are formed in the transport of eight molecules of acetyl CoA to the cytoplasm. The additional six NADPH needed come from PPP.
Acetyl CoA carboxylase in Controlling Fatt Acid Metabolism
Fatty acid synthesis is maximal when carbohydrates and energy are plentiful and when fatty acids are scarce. Recall, acetyl CoA carboxylase catalyzes the committed step in fatty acid synthesis (the conversion of acetyl CoA to malonyl CoA), and thus is an important regulatory enzyme of fatty acid metabolism.
Acetyl CoA carboxylase is switched off by phosphorylation and activated by dephosphorylation. AMP-dependent protein kinase converted acetyl CoA carboxylase into an inactive form by modifying a serine residue. AMP-dependent kinase is activated by AMP signifying that energy is low and synthesis must stop.
The carboxylase is also allosterically stimulated by citrate. Citrate facilitates the polymerization of the dimers of the carboxylase, thus partially reversing the inhibition caused by phosphorylation. Citrate levels rise both when acetyl CoA and ATP are abundant, signifying that energy is available for fatty acid synthesis.
Malonyl CoA also has a role in regulation. Malonyl CoA is abundant when fuel molecules are abundant. Malonyl CoA inhibits carnitine acyltransferase I, preventing the entry of fatty acyls into the mitochondria for degradation.
Glucagon and epinephrine inhibit fatty acid synthesis, although the exact mechanism is unknown. But basically, glucagon signals that fuel is low and epinephrine signals the immediate need for energy, which enhances the glucagon effect.
Insulin will acitvate fatty acid synthesis by stimulating the activity of a protein phosphatase.